Article Text

Abstract
BACKGROUND AND AIMS The main goal of our study was to characterise the activity of BIM26226 as a peripheral gastrin releasing peptide (GRP) receptor antagonist in healthy human subjects and to determine if endogenous GRP is a physiological regulator of gastric acid secretion and gastrin release.
METHODS Our study consisted of three parts. In part I, subjects received saline or BIM26226 followed by graded doses of intravenous human GRP in a four period crossover design. In part II, subjects received BIM26226 or saline during oral meal ingestion or modified sham feeding. In part III, subjects received an acidified meal in the presence and absence of BIM26226 in a two period crossover design. In addition, gastrin and somatostatin mRNA were measured in biopsy specimens during saline and BIM26226 infusion.
RESULTS BIM26226 dose dependently inhibited GRP induced acid output. Acid secretion after oral liquid meal intake and sham feeding was significantly inhibited by BIM26226 (p<0.01) whereas plasma gastrin release remained unchanged. Gastrin and somatostatin mRNAs were not significantly different after saline or BIM26226.
CONCLUSIONS BIM26226 is a potent GRP antagonist in humans. Endogenous GRP may be a physiological regulator of gastric acid secretion. Gastrin release does not seem to be under the control of GRP.
- gastrin
- gastric acid secretion
- gastrin releasing peptide
- gastric function
- BIM26226
Abbreviations used in this paper
- GRP
- gastrin releasing peptide
- PEG
- polyethylene glycol
- CCK
- cholecystokinin
- GAPDH
- glyceraldehyde phosphate dehydrogenase
- NMB
- neuromedin B
- ECL
- enterochromaffin-like
Statistics from Altmetric.com
In the human gastrointestinal tract, bombesin-like immunoreactivity is mainly found in the stomach and pancreas.1 ,2 Infusion of bombesin dose dependently increases plasma gastrin and gastric acid secretion. A maximal acid response is seen with low doses of bombesin whereas plasma gastrin is further stimulated with increasing doses. Compared with stimulation with pentagastrin, the acid response to bombesin-like peptides is clearly submaximal. Failure of bombesin to increase both effects in parallel is in line with the hypothesis that an unknown inhibitor of acid secretion is released at the same time.3 ,4 Current data indicate that stimulation of acid secretion predominantly occurs in a paracrine/neurocrine manner (histamine, acetylcholine) whereas the stimulating effect mediated by gastrin may be less important.5
In humans, gastrin releasing peptide (GRP) seems to stimulate acid secretion by release of gastrin.6 ,7 Studies measuring the effect of intravenous GRP infusion have demonstrated that low, so called physiological, concentrations of GRP stimulate gastrin release and acid secretion in healthy volunteers. However, the physiological relevance of these observations is limited by the fact that GRP is a neuropeptide and not a circulating hormone. To date, it has not been possible in humans to selectively inhibit GRP release from nerve terminals or to block the action of GRP by inhibiting its binding in target tissues.
Recently, [D-F5-Phe6,D-Ala11]Bn(6–13)OMe (BIM26226) has been developed which is specific for GRP preferring bombesin receptors.8 In rat pancreatic acini as well as in the rat pancreatic carcinoma cell line AR4-2J, BIM26226 had the same binding affinity as bombesin and GRP but at least 300-fold lower affinity for neuromedin B (NMB) preferring bombesin receptors.9 BIM26226 potently inhibited bombesin stimulated amylase release from both rat pancreatic acini and AR4-2J cells. Intravenous bolus injection inhibited bombesin stimulated amylase release for up to 4.5 hours,10 suggesting that in animals, BIM26226 is a potent and specific GRP antagonist in vitro and in vivo.
The present investigation comprised three parts (I–III). In part I, subjects received saline (control) or one of three intravenous doses of BIM26226 followed by graded doses of intravenous human GRP in a four period crossover design. The objectives of this part of the study were to demonstrate that intravenous BIM26226 is an effective GRP receptor antagonist in humans and to establish the dose range of antagonist activity based on inhibition of GRP stimulated acid secretion and hormone release. In part II, subjects received a continuous infusion of BIM26226 or saline (control) followed by oral ingestion of a meal or modified sham feeding. The aim of this part of the study was to assess the physiological role of endogenous GRP on meal induced intragastric acidity, sham feeding stimulated acid output, and meal stimulated plasma gastrin response. In part III, subjects received an acidified meal in the presence and absence of BIM26226 in a two period crossover design. The objective was to investigate plasma gastrin release under GRP receptor blockade at controlled intragastric pH.
Methods
EXPERIMENTAL SUBJECTS
Thirty healthy male subjects, 20–48 years of age, participated in the studies. Each subject gave written informed consent and had a normal physical examination and laboratory tests. All subjects wereHelicobacter pylori negative (13C urea breath test). The study was approved by the ethics committee of the University Hospital of Basel.
BIM26226
BIM26226 was synthesised by DH Coy at Tulane University, New Orleans, Louisiana, USA (chemical purity >97%) and provided as lyophilised powder by Biomeasure Inc. (Milford, Massachusetts, USA). In addition to GRP/NMB receptors, BIM26226 has been examined for its in vitro binding to a total of 26 different receptors, among them cholecystokinin (CCK)A/B, peptide YY1/2, somatostatin2/5, histamine2, PACAP1/2, secretin, and vasoactive intestinal peptide. Specificity of binding was demonstrated by Ki values >1000 nM for all non-bombesin receptors tested (John Taylor, personal communication). Plasma concentrations of BIM26226 at an infusion rate of 500 μg/kg/h were in the range 200–400 nmol/l whereas doses of 50 and 5 μg/kg/h induced 10- and 100-fold lower concentrations. Fifteen minutes after the end of infusion, the compound was not detectable, indicating a plasma half life of less than two minutes. Based on plasma concentrations induced by the highest dose of BIM26226, it can be estimated that the ratio of agonist/antagonist at the receptor site is in the range 1:1000, which is necessary to inhibit function by more than 97%.11
STUDY PROTOCOLS
Each experiment was performed on a different day and in random order. Subjects were fasted overnight before each experiment.
Effect of BIM26226 on GRP stimulated acid output
Eight subjects participated in a four period, double blind, placebo controlled, crossover study. On each experimental day subjects swallowed a double lumen gastric tube which was positioned under fluoroscopic control. On four days, separated by at least three days, volunteers received intravenous infusions of saline (control) or one dose of BIM26226 (5, 50, or 500 μg/kg/h). After a baseline period of 60 minutes, increasing doses of synthetic human GRP (10, 30, and 90 pmol/kg/h; Novabiochem, CH-4448 Läufelfingen, Switzerland) were given intravenously, each dose for 45 minutes. Continuous intragastric perfusion of a non-absorbable marker, polyethylene glycol (PEG4000), was administered throughout the study at a rate of 3 ml/min to correct for duodenal loss. Gastric acid secretion was collected continuously in 15 minute aliquots. Blood was drawn at various time points for determination of plasma gastrin.
Effect of BIM26226 on meal stimulated intragastric acidity and plasma gastrin release
In 10 subjects intragastric acidity was measured by miniature combined glass electrodes (Ingold Lot 440; CH-8902 Urdorf, Switzerland) placed in the gastric body and connected to a portable pH data logger (gastrograph Mark III; Medical Instruments Corp., CH-4500 Solothurn, Switzerland). The electrodes were calibrated at 20°C using commercial buffer solutions, and temperature correction for intragastric readings (37°C) was performed. The electrodes were passed transnasally and positioned 8–10 cm below the lower oesophageal sphincter; the correct position was verified by fluoroscopy. Intragastric pH was measured every six seconds throughout the experiments. On two different days, subjects received intravenous infusions of either saline (control) or BIM26226 (500 μg/kg/h) in a double blind crossover manner. After a baseline period of 120 minutes, each subject ingested a liquid 500 kcal test meal (500 ml Ensure; Abbott, CH-6330 Cham, Switzerland) within 10 minutes while pH monitoring continued. After four hours, a second 500 kcal Ensure meal was ingested and pH monitoring continued for another three hours. Blood was drawn at various time points for measurement of plasma gastrin.
Effect of BIM26226 on modified sham feeding stimulated acid output
In six subjects a double lumen gastric tube was inserted transnasally into the stomach on two different days. They received a meal consisting of sandwiches (cooked ham, white bread and butter) and 200 ml of milk for modified sham feeding using the chew and spit technique.12
Volunteers were trained in advance not to swallow any food, liquid, or saliva. Recovered gastric juice was visually monitored and showed no evidence of accidentally swallowed food. Gastric acid secretion was continuously quantified in 15 minute aliquots as described above. PEG4000 was used as a non-absorbable marker to correct for duodenal loss. The experiments were performed with intravenous saline infusion (control) or with BIM26226 (500 μg/kg/h) in a double blind crossover fashion. Saline or BIM26226 was started 60 minutes before modified sham feeding. Blood was drawn as described above.
Effect of BIM26226 on plasma gastrin at constant intragastric pH
Six subjects swallowed a pH electrode on two different days. After a two hour baseline period, an acidified liquid test meal (Ensure) was perfused into the stomach at a rate of 8 ml/min over 60 minutes. The meal was prepared by mixing 60 ml of 1 M HCl thoroughly with 500 ml of Ensure, giving a pH of 2.3–2.5. The experiments were performed twice in each subject, once with intravenous saline infusion (control) and once with BIM26226 (500 μg/kg/h) in a double blind crossover design. Blood was drawn at regular intervals for determination of plasma gastrin.
Effect of BIM26226 on gastrin and somatostatin mRNAs
Seven subjects received either intravenous saline or BIM26226 (500 μg/kg/h) for three hours on separate days. At the end of the infusion, they underwent gastric endoscopy. Mucosal biopsies (four from the antrum and two from the corpus) were taken and immediately stored in liquid nitrogen until further analysis. Quantitative determination of gastrin and somatostatin mRNA compared with glyceraldehyde phosphate dehydrogenase (GAPDH) mRNA, extracted from gastric biopsies, was performed by northern blotting as described previously.13
EXPERIMENTAL MEASUREMENTS
Gastric acid secretion
The volume of all gastric aspirates was measured to the nearest 1 ml and gastric juice was analysed for [H+] and [PEG4000]. Acid concentration was determined by the backtitration method and PEG4000 turbidimetrically.14 The marker perfused in the stomach served as a control parameter to calculate acid output for a given period.15 Results are expressed as the mean of the last two 15 minute periods during each dose. Data are presented as mean (SEM). Recovery rate was 86 (7)%.
Gastrin radioimmunoassay
Plasma gastrin concentration was measured using a commercially available antiserum with negligible cross reactivity with CCK or other gastrointestinal peptides. The sensitivity of the assay was 25 pg/ml. All samples from a single protocol were run in the same assay with an intra- and interassay variation of <10% (Pantex, Santa Monica, USA).
STATISTICAL ANALYSIS
Data were calculated using an analysis of variance model for either a four or two period crossover design followed by a parametric or non-parametric test, as appropriate.16 Data were tested for normal distribution using the Komolgorov-Smirnov test and for homogeneity of variances according to Levine's test.16Intragastric acidity readings were transferred from the portable registration unit to a computer and mean pH values for one minute intervals were calculated. From individual one minute pH values, medians for the fasting state and after meal intake were calculated. Group medians were compared using the Wilcoxon signed rank test. Gastrin results are expressed as mean (SEM). Integrated plasma hormone and gastric acid responses are expressed as area under the output curve, defined as the increase over basal values and calculated according to the trapezoid rule.16
Results
EFFECT OF BIM26226 ON GRP STIMULATED ACID OUTPUT AND PLASMA GASTRIN RELEASE
Subjects received intravenous infusions of saline (control) or one dose (5, 50, or 500 μg/kg/h) of BIM26226. After a baseline period of 60 minutes, graded doses of human synthetic GRP (10, 30, and 90 pmol/kg/h) were administered, each dose given for 45 minutes. Basal acid output in the control experiments was 1.27 (0.24) mmol/15 minutes. The lower two doses of BIM26226 had no effect (1.35 (0.35) and 1.11 (0.12) mmol/15 minutes, respectively) whereas 500 μg/kg/h significantly inhibited basal acid output (0.47 (0.06) mmol/15 minutes; p<0.05). Changes in acid output in response to graded GRP infusion and the ability of various doses of BIM26226 to affect this change are shown in fig 1A. In the saline (control) experiment, infusion of GRP promptly stimulated gastric acid secretion with a maximal secretory response obtained at 30 pmol/kg/h. The highest dose of GRP (90 pmol/kg/h) failed to further increase acid output although plasma gastrin values continued to rise (see below). In comparison with stimulation with pentagastrin, the response to exogenous GRP infusion was clearly submaximal.

(A) Effect of saline or one dose of BIM26226 (5, 50, 500 μg/kg/h) on gastric acid output during infusion of graded doses of synthetic human gastrin releasing peptide (GRP) (0, 10, 30, 90 pmol/kg/h). Acid output is presented as mmol/15 minutes and results are mean (SEM) of eight subjects. BIM26226 dose dependently inhibited integrated acid responses; a dose of 500 μg/kg/h completely blocked basal and maximally GRP stimulated acid secretion (p<0.01). (B) Plasma gastrin concentrations during the same study. Results are expressed as mean (SEM). BIM26226 dose dependently inhibited integrated gastrin responses; 500 μg/kg/h completely blocked GRP stimulated plasma gastrin release (p<0.01).
BIM26226 dose dependently inhibited GRP induced acid output. Although 5 μg/kg/h BIM26226 did not significantly modify the integrated response to GRP, the dose response curve was shifted to the right with the same maximal response. A dose of 50 μg/kg/h significantly decreased the integrated acid response compared with control values (p<0.05), and 500 μg/kg/h BIM26226 completely inhibited GRP stimulated acid secretion in the dose range tested (p<0.01v placebo). Mean basal gastrin concentrations were not different between the four treatments. Graded doses of GRP induced increasing plasma gastrin concentrations in the saline (control) experiment which were dose dependently inhibited by increasing doses of BIM26226 (fig 1B).
EFFECT OF BIM26226 ON MEAL STIMULATED INTRAGASTRIC ACIDITY AND PLASMA GASTRIN
Subjects received either intravenous saline (control) or BIM26226 (500 μg/kg/h). The 11 hour pH profiles of both treatments (placebo or BIM26226) are shown in fig 2A. Median pH (interquartile range) in the pre-lunch period (0–2 hours) was 1.2 (range 1.0–1.5) with placebo. With BIM26226, median pH was 1.8 (range 1.5–2.0; p<0.05). In the placebo (saline) experiments, intragastric acidity after both meals showed a similar pattern; pH increased to 4.5 and returned to basal values within three hours. In contrast, with BIM26226 postprandial pH was 4–6 after the first meal and remained elevated during the whole interdigestive period. The second meal did not induce any further changes in pH. Total 11 hour intragastric pH was significantly higher with BIM26226 (4.1 (0.5)) compared with placebo (1.4 (0.4); p<0.01). We cannot exclude the fact that the major effect of the antagonist was primarily on basal acid secretion; however, the study with cephalic stimulation of acid secretion favours the interpretation that BIM26226 inhibited basal and stimulated acid output (see below).
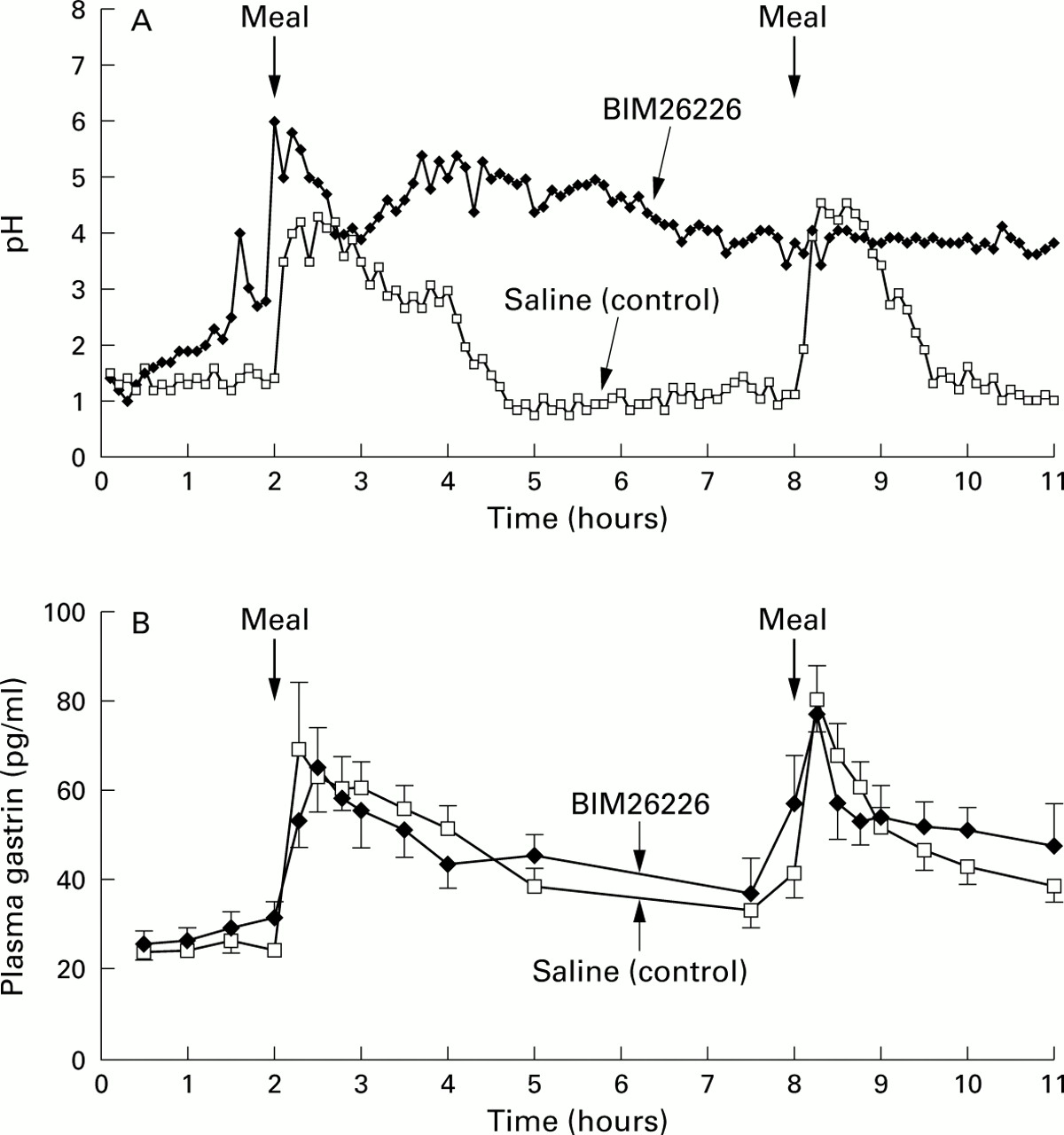
(A) Time course of intragastric acidity before and after oral ingestion of two liquid meals with and without concomitant infusion of BIM26226 (500 μg/kg/h) in 10 healthy volunteers. Data are presented as medians. Intragastric acidity was significantly increased during infusion of BIM26226 (p<0.01). (B) Plasma gastrin concentrations during the same study with and without BIM26226 (500 μg/kg/h). Results are expressed as mean (SEM). Basal and postprandial plasma gastrin levels were not significantly different with and without BIM26226.
Plasma gastrin levels before and after meal intake are shown in fig 2B. Pre-lunch (0–2 hours) levels were not significantly different although values during BIM26226 were slightly higher. The first meal induced a rapid increase with a subsequent decrease during the postprandial period (2–5 hours). A similar pattern was seen after the second meal. BIM26226 did not suppress meal stimulated gastrin release.
Modified sham feeding resulted in a significant increase in acid output which was suppressed by intravenous BIM26226 (fig 3A). In the control study, plasma gastrin concentrations were increased in response to modified sham feeding but the difference compared with basal levels was not statistically significant. The lack of a significant increase in gastrin levels could have been caused by a type II error, as only a small number of subjects were tested. Plasma gastrin concentrations before and after sham feeding were not statistically different with and without BIM 26226 (fig 3B).
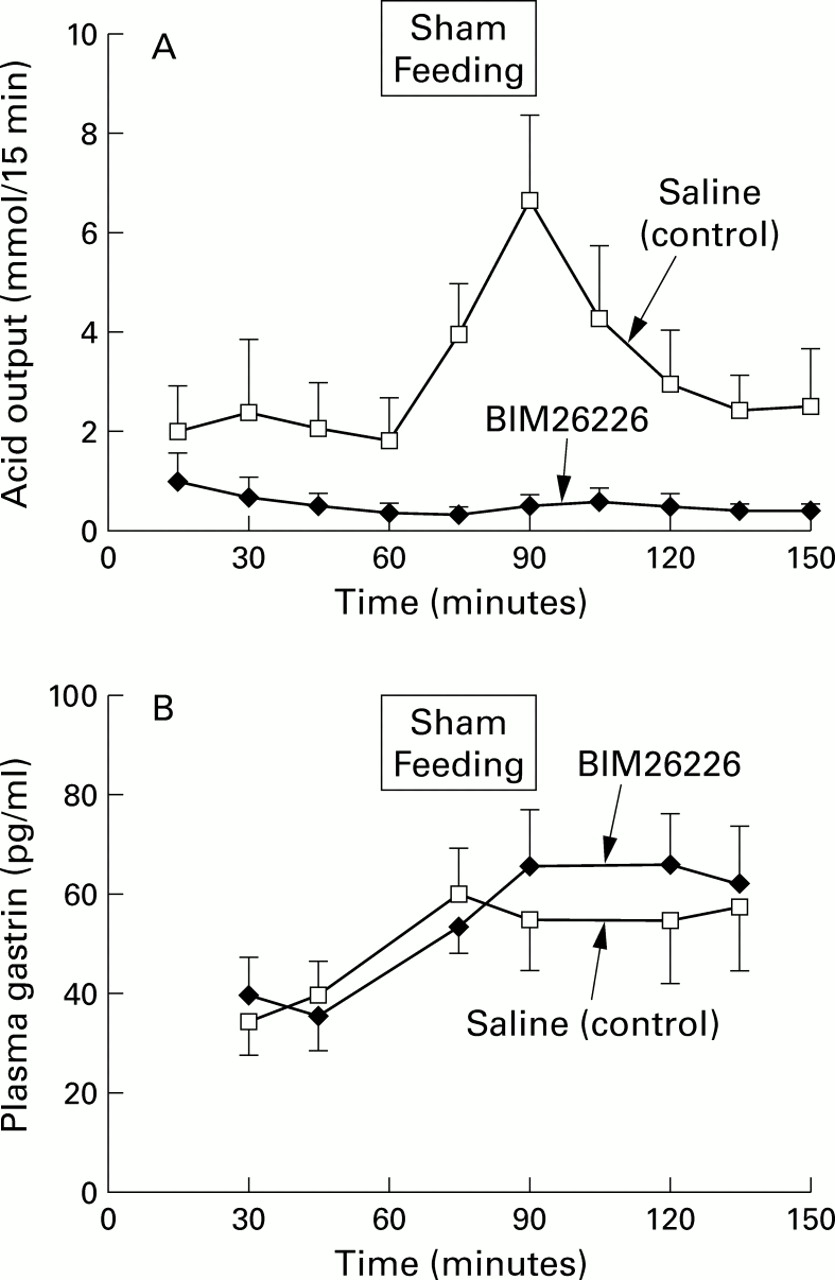
(A) Gastric acid output (mmol/15 minutes) in response to modified sham feeding with and without concomitant BIM26226 (500 μg/kg/h) in six healthy subjects. Data are presented as mean (SEM). BIM26226 significantly suppressed acid output compared with control values (p<0.05). (B) Plasma gastrin concentrations during the same study. Results are expressed as mean (SEM). Plasma gastrin levels before and after sham feeding were not significantly different with and without BIM26226.
EFFECT OF BIM26226 ON PLASMA GASTRIN AT CONSTANTLY LOW INTRAGASTRIC pH
Perfusion of an acidified liquid test meal at pH 2.3 resulted in a steady intragastric pH. In each single experiment, for both intravenous saline and BIM26226 infusion, median intragastric pH was below 2.3 (BIM26226 median pH 1.7 (range 1.4–2.2); saline median pH 1.3 (range 1.0–1.7)). Perfusion of the meal into the stomach induced a significant increase in plasma gastrin (p<0.05) whereas BIM26226 did not affect this response (fig 4).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Plasma gastrin responses before, during, and after intragastric instillation of an acidified liquid meal with and without concomitant intravenous BIM26226 in six healthy subjects. Basal as well as meal stimulated gastrin release were not significantly changed by BIM26226.
EFFECT OF BIM26226 ON GASTRIN AND SOMATOSTATIN mRNA
Biopsy specimens taken at the end of three hours of saline or BIM26226 infusion were quantified for gastrin mRNA, somatostatin mRNA, and GAPDH mRNA by northern blot analysis. Results are expressed as a ratio of peptide mRNA to GAPDH mRNA (mean (SEM), n=7). Gastrin mRNA to GAPDH mRNA ratio in antral samples was not different with (1.66 (0.39)) or without (1.91 (0.37)) BIM26226. Ratios of somatostatin mRNA to GAPDH mRNA were not altered with or without BIM26226, either in the antrum (0.179 (0.042) v 0.199 (0.038)) or the corpus (0.094 (0.021) v 0.068 (0.016)).
ADVERSE EFFECTS
No adverse effects or significant changes in blood chemistry, urine, or electrocardiograms were observed after intravenous administration of BIM26226.
Discussion
In our study, we showed for the first time that BIM26226 is a potent GRP receptor antagonist in humans. BIM26226 inhibited exogenously GRP stimulated gastric acid output and plasma gastrin release dose dependently. Furthermore, BIM26226 markedly inhibited intragastric acidity and completely suppressed sham feeding stimulated acid output but had no effect on plasma gastrin in response to either food or sham feeding. Based on our data, we propose that GRP is a physiological regulator of gastric acid secretion. Gastrin release however does not seem to be a physiologically relevant GRP mediated effect.
Gastric acid secretion is regulated by a complex interaction of neural, paracrine, and endocrine signals from the brain and gastrointestinal tract. GRP has been associated with meal stimulated acid stimulation by gastrin release. Regulation of gastrin secretion includes stimulatory factors such as acetylcholine and local luminal factors, including protein digestion products and luminal acidity17 but also inhibitory pathways such as somatostatin.5 ,18 ,19
The main finding of our study was that acid secretion after oral liquid meal intake and sham feeding was markedly inhibited by GRP receptor blockade whereas plasma gastrin release remained unchanged. Several studies have shown that intravenous GRP/bombesin stimulate gastrin release and acid output in humans.4 ,6 ,7 ,20 This effect however may be more of a pharmacological than physiological event. The dose dependent inhibition of GRP induced acid secretion in our study demonstrates the antagonistic activity of BIM26226 in humans which has previously been shown only in animals.9 ,10 The ability of BIM26226 to completely block acid output in response to modified sham feeding and to markedly suppress intragastric acidity after oral food indicates that endogenous GRP is involved in the regulation of postprandial acid production in humans. This action is not mediated by gastrin. The observation is further supported by the finding of an unchanged gastrin mRNA. Furthermore, somatostatin mRNA was not altered by the GRP receptor antagonist. The finding is at odds with the hypothesis that elevated somatostatin mRNA expression in the antrum mucosa (acting on G cells) or in the corpus (directly inhibiting the parietal cell) could have explained BIM26226 mediated inhibition of acid secretion.
Based on our findings, we tried to define the potential role of GRP in regulating gastric acid secretion. Stimulation of the vagus nerve induces acetylcholine release near parietal cells, histamine containing cells, and somatostatin producing D cells.21-23Muscarinic receptors of the M3 type have been identified on parietal cells24 but muscarinic receptors are probably also present on ECL cells and on D cells. Stimulation of acetylcholine release activates several stimulatory pathways of acid production, such as direct stimulation of the parietal cell and indirect stimulation of the parietal cell through stimulation of histamine release by enterochromaffin-like (ECL) cells. Histamine stimulates parietal cell acid production through activation of histamine receptors.25 Muscarinic receptors on D cells potentially inhibit somatostatin release thereby reducing the inhibitory effect of somatostatin on ECL and parietal cells. We suggest that GRP acts from an intramural ganglion within the N vagus to regulate acetylcholine release but this last statement requires experimental confirmation. Thus increased acetylcholine secretion results in direct stimulation of the parietal cell and in histamine release from ECL cells. As histamine and acetylcholine interact at the level of the parietal cell to stimulate acid secretion, interference with these two stimulatory pathways through endogenous blockade could induce a marked reduction in acid output. We believe that GRP is a regulator of acid production in humans but the hypothesis requires experimental support.
The physiological role of endogenous GRP in regulating gastrin secretion in humans remains unknown. Our results have clearly shown that GRP can regulate gastric acid secretion independent of gastrin release. Several aspects remain unclear. Firstly, GRP preferring bombesin receptors on human G cells have not been investigated in detail. Secondly, bombesin and GRP enhanced gastrin release from cultured human G cells but the action of a putative bombesin antagonist was difficult to assess.24 ,26 Furthermore, species differences in animal models make it difficult to extrapolate findings of these studies to humans. As gastrin release is regulated by various other factors, including nutrient components, alteration in gastrin secretion with BIM26226 may be demonstrable under other experimental conditions.
In conclusion, we have demonstrated that BIM26226 is an effective GRP receptor antagonist with respect to inhibition of GRP stimulated gastrin release and acid production in humans. Furthermore, these studies have confirmed the usefulness of BIM2662 as a tool to investigate the physiological activity of endogenous GRP, suggesting a role for this neuropeptide in regulating acid secretion in humans.
Acknowledgments
We thank Carita Frei for editorial assistance and for preparation of the manuscript. The support of the Swiss National Science Foundation (grant No 3200-40604.94/1) and Ipsen International, London, UK, is gratefully acknowledged.
Abbreviations used in this paper
- GRP
- gastrin releasing peptide
- PEG
- polyethylene glycol
- CCK
- cholecystokinin
- GAPDH
- glyceraldehyde phosphate dehydrogenase
- NMB
- neuromedin B
- ECL
- enterochromaffin-like
References
Linked Articles
- Commentary
- LETTER